Search for Pathogens
Q: How Does the Immune System Quickly Recognize Foreign Things and Become (Long-Term) Tolerant to non-Pathogenic Material
The immune system is fundamentally an information system. Information comes into the system in the form of proteins and other chemicals and cues. Each cell of the immune system is its own 'agent' to sense one or more of these and each also gets to give and receive cues from other cells. T cells get their primary cues in the forms of peptide 'antigens' that are presented to them by phagocytes or other cells that have already encountered the cue (virus, bacteria etc.) Ultimately, for T cells to get these cues, some cell needs to bear the peptide and present it. And T cells need to then survey those cells and, using their T cell receptors, recognize the antigen and begin to react.
Current Project: The Distribution of Antigens
In the past six years, we've begun to focus on the antigen-presenting cell as the cell that ultimately controls what a T cell can respond to. In effect, the logic of the immune system--having one cell be required to present to another--forces T cells to be subservient to the directives of these antigen-presenting cells. An important clue seems to be that antigen-presenting cells 'hold' antigens in a vesicular compartment for long periods of time--perhaps a history of their exposure. They then can either present that material to T cells, degrade it, or hand the vesicle off to again make those same decisions. A diagram of a bit of this process is from our 2016 Cancer Cell paper:
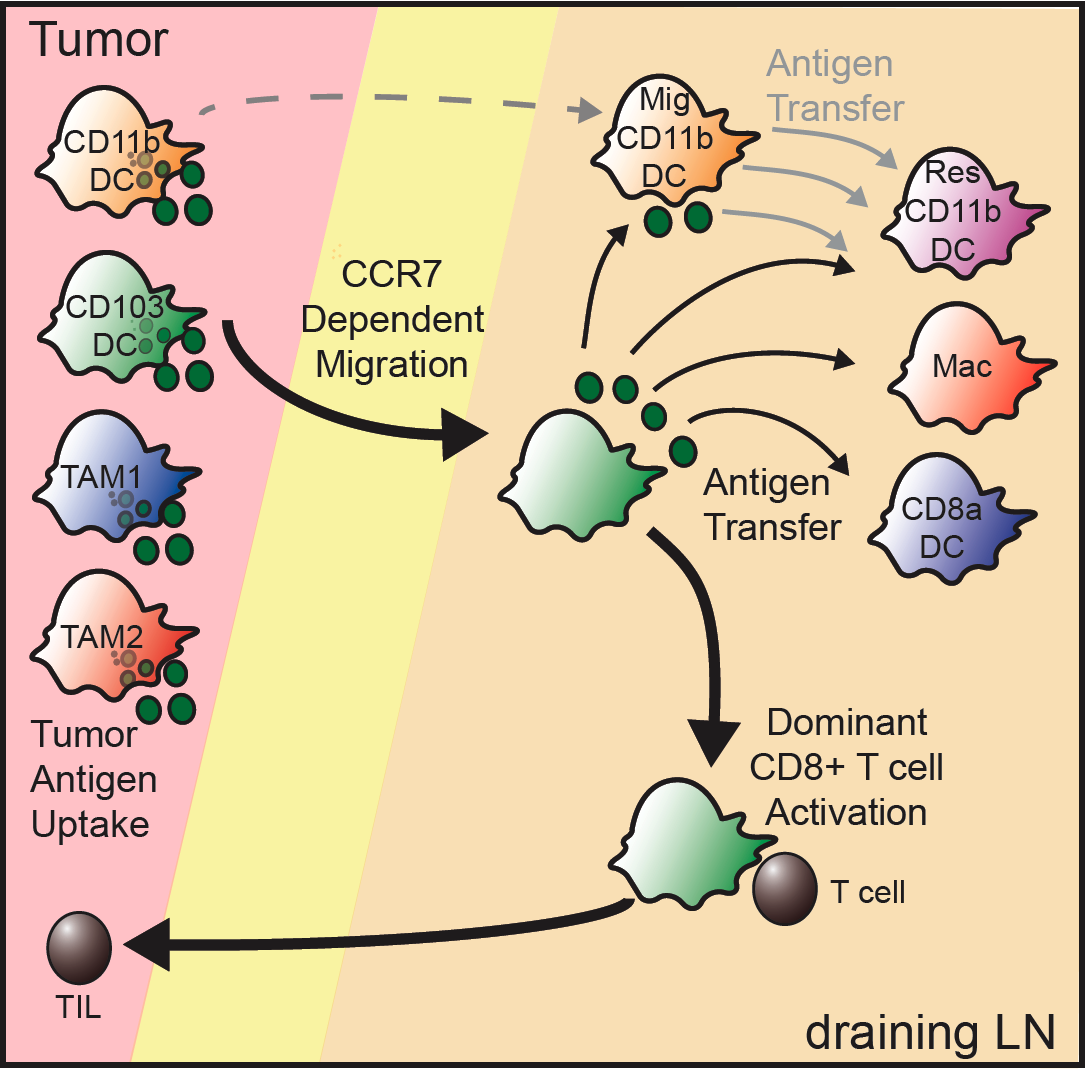
So what are these cell types, what does each one uniquely do when it interacts with T cells. We are currently working to describe the entirety of this process--from the kinetics to how vesicles are 'encoded' for their contents, be it self, tumor, commensal or pathogen. We seek to understand how each antigen-presenting cell type holds antigens and how might we 'hack' the system of antigens in order to influence T cell immunity.
Historical: The Search Process
We have been interested in the fundamental behaviors that cells use to 'see' their world. A big component of this is how T cells move through tissues to 'search' for evidence of their antigens. A big component of that is governed by the motility machinery in T cells. In studying a myosin1g loss-of-function T cells,(Gerard et al. 2014) we identified this motor as a ‘steering’ motor in cells that forces them to make periodic turns. Without the motor, T cells crawl more quickly but in straighter paths. We showed that this rapid and straight movement through a tissue is ultimately bad for search--T cells that lacked this motor protein could be stimulated fine when we gave them lots of antigen presenting cells but when antigen presenting cells were in limited supply, T cells often missed engaging them. This meant that animals with T cells lacking this protein, exposed to a low dose of material from a pathogen, were not protected (vaccinated) as were normal mice. 'Search' is important!
Our first entry into this field when we proposed that motile forces and their control underlie the control of immunological synpases (IS) and similar synapses between immune cells—sites where information exchange is enhanced. We thus focused on candidate myosin motors that are expressed in T cells. Our lab was the first to identify and characterize the major Myosin II isoform in T cells. In Jacobelli et al. Nature Immunology, 2004, we showed MyosinIIA (“MyoIIA”. from the gene MyH9) was the only isoform expressed in T cells and was necessary for effective motility in vitro in a non-confined environment. In this setting, myosinII contractility was associated with stable motile propagation as well as generation of cortical tension associated with the amoeboid shape of motile T cells in such non-confined environments. We also showed TCR and Ca-triggered phosphorylation of the MyoIIA protein, consistent with a transient inactivation of this motor during the generation of the immunological synapse (IS). We now hypothesize that this ‘de-activation’ is a critical part of the ability of T cells to quickly ‘spread’ onto antigen presenting cells and set up an effective IS.
In Jacobelli et al. J. Immunology, 2009, we demonstrated that this motor plays a critical role in determining how cells approach a substrate, especially under confinement. This study used TIRF imaging to demonstrate two unique ‘modes’ of amoeboid movement. In the first, which we term ‘walking’, T cells utilize MyosinIIA to generate multiple ‘footprints’ on substrates. Typically as MyosinIIA contracts inward and extinguishes one footprint, another adhesion is licensed nearer the leading edge. This mode is quite distinct from what we term ‘sliding’, which more closely resembles what is observed for mesenchymal motility. Under sliding, which does not require MyosinIIA, cells maintain a continuous and much larger adhesion with substrate. This mode is slower than ‘walking’ but also likely permits more effective surface scanning and we thus hypothesize that inactivation of MyoIIA at the start of IS formation (via phosphorylation as described in Jacobelli, 2004) is likely critical to increasing sensitivity to pMHC complexes.
In Jacobelli et al. Nature Immunology, 2010, we sought to analyze the full import of MyosinIIA expression in T cells in complex 3-dimensional environments. By generation of mice in which MyoIIA was specifically deleted in naïve T cells or activated T cells derived from them, we demonstrated that these cells showed profound overadhesion during motility, resulting in their slowed movement as they scan through lymph nodes. To analyze the membrane dynamics underlyin this, in greater detail, we generated artificial 3-dimensional environments via microfabrication techniques. Beyond recapitulating the overadhesion/slowing in lymph nodes, we found that T cell migration rate is regulated by the degree to which the cells are confined rather than merely by the stickiness of the surfaces along which they crawl as had previously been thought (the ‘haptokinetic model’). For amoeboid cells, we found a confinement-optimized ‘channel size’ for optimal motility rate that is just a bit wider than the cells average width. We hypothesize that when cells are further squeezed, they slow down due to increased friction with the substrate walls. In contrast when they are confined less, they appear not to be able to efficiently generate the multiple footprints that defined the ‘walking’ mode of our previous studies and that characterized the optimum movement. This ability to optimize speed in turn was reliant on the presence of MyosinIIA to regulate the adhesions made during ‘walking’.